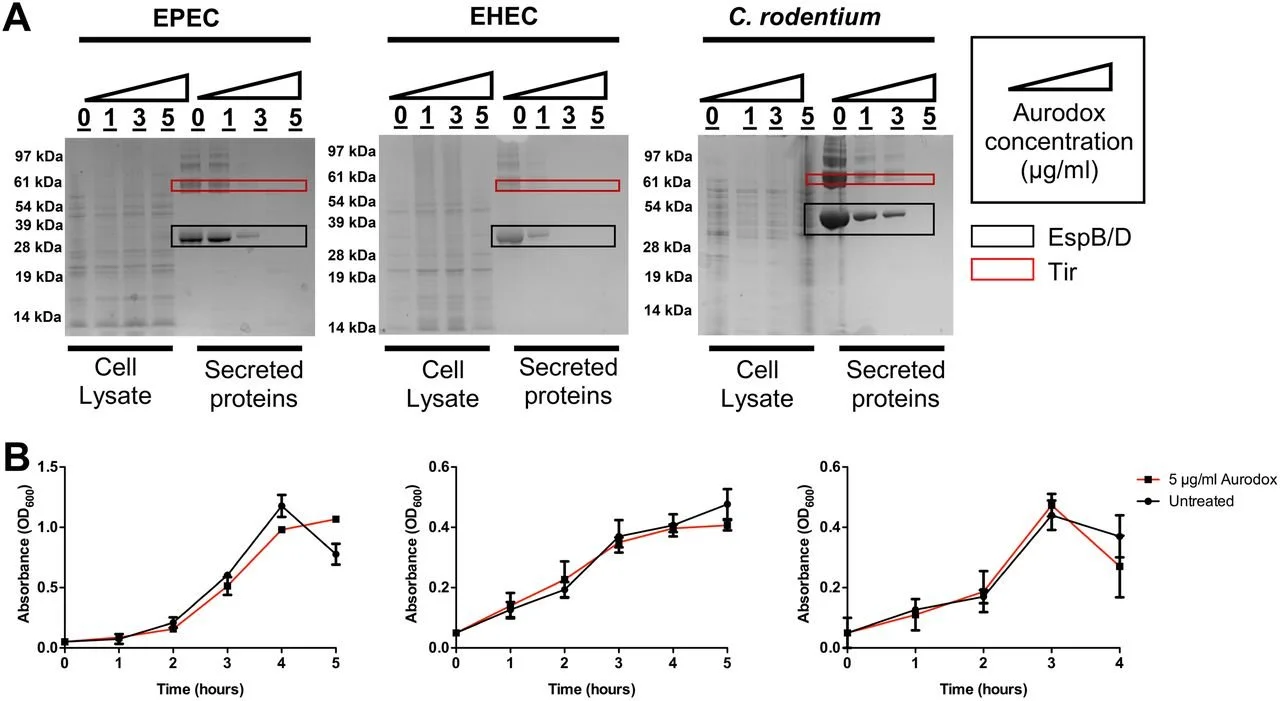
by Esha Ghai
We have long been told that our DNA is unchangeable, often encouraging us to blame our flaws and accredit our talents to genetics. However, the introduction of genome-editing gives scientists the ability to alter genetic material and effectively change the basis of an organism. Genetic material can now be added, removed, or mutated at any specified location in an organism, allowing for cures to genetically inherited diseases and a wide range of illnesses. A recently discovered microbial ribonucleoprotein with such genome-editing capabilities is the CRISPR/Cas9 system.
CRISPR/Cas9, short for “clustered regularly interspaced short palindromic repeats and CRISPR-associated protein 9”, was naturally developed by bacteria as a defense mechanism against invading viruses. As a part of this mechanism, bacteria confiscate fragments of DNA from invading viruses, and use the fragments to construct CRISPR arrays [1]. CRISPR arrays give the bacteria the ability to recognize identical or similar viruses, so if a similar virus attacks again, the bacteria produces RNA segments from the CRISPR array to specifically attack the virus’s DNA [1]. Cas9, an enzyme, is then used to cut apart the DNA, essentially killing the virus [1]. Scientists in the medical community are now adapting this gene-manipulation system to be used in cells in other organisms, including humans. The genome-editing tool has unlocked the potential to combat human genetic diseases previously deemed untreatable. A more cost-efficient, advanced, and precise method of gene-editing, CRISPR/Cas9 is revolutionizing the face of medicine as we know it [1].
Fascinated by the properties of CRISPR/Cas9 and microbes, Dr. Xue Gao, a professor of Chemical and Biomolecular Engineering at Rice University, currently investigates the applications of genome-editing and microbe-based natural product discovery. Her research “lies at the interface of chemical biology and biomolecular engineering with a primary focus on small- and macro-molecule discovery and their applications to human health, agriculture, and energy.” Dr. Gao’s contribution to the development of Statins, a pharmaceutical drug with lipid-lowering capabilities for those at risk of cardiovascular disease, piqued her interest in protein engineering. The frequent use of Statins in the medical society motivated Dr. Gao to continue synthesizing molecules to combat diseases.
Her research also encompasses the use of enzyme-catalyzation (increase in the rate of a process due to an enzyme) for anticancer and antibiotic drug synthesis. Enzymes are powerful biological catalysts, accelerating chemical reactions by assisting in the formation of a transition state between the starting reagent and end product. Enzymes are also highly specific in which reactions it catalyzes, usually catalyzing only a single chemical reaction (or reactions very similar in nature), limiting side reactions and the ultimate formation of wasteful by-products [3]. According to Dr. Gao, whereas traditional chemical methods of synthesizing compounds require environmentally toxic solvents and harsh conditions, enzymes work well in moderate environments and are capable of producing a higher yield of 99%. As enzymes are proteins, extreme temperatures and pH levels result in denaturation, rendering the enzyme inactive. Therefore, enzyme catalyzation ability is enhanced at moderate conditions. In biological systems, a majority of processes utilize enzymes, from the transfer of CO2 in the body through the enzyme carbonic anhydrase, to breaking down sugars through the enzyme amylase. The development of enzymes to catalyze complex chemical reactions, called directed evolution, is a major focus of the Gao Laboratory.
Dr. Gao ascribes “her love [for] exploring what has already been designed by nature” to the sheer intelligence of microbes and their methods of defense against viruses. “Microbes are super smart. They create protein encodings themselves to fight different types of environments.” CRISPR/Cas9, another prominent example utilizing protein encodings and enzyme catalysis, has also attracted Dr. Gao’s interest. CRISPRs are specialized protein encodings, while Cas9 is an enzyme capable of cutting apart genetic material. Recently, Dr. Gao utilized the genome-editing capabilities of CRISPR/Cas9 to treat autosomal-dominant hearing loss. Autosomal-dominant diseases are inherited from an abnormal gene from one parent, even when the corresponding gene from the other parent is normal. Dr. Gao’s findings suggested that a targeted delivery of gene-disrupting agents such as CRISPR/Cas9 in vivo (within a living organism) can potentially treat such cases of hearing loss [2].
Dr. Gao’s research group is planning to do further work in CRISPR/Cas9 applications for a variety of human genetic diseases, and hopes to make breakthroughs in research through accommodating advanced computational tools and developing DNA sequencing technology (technology that determines the sequence of nucleotide bases). Dr. Gao envisions the next stage of her group’s research to be developing new enzyme-catalyzed reactions in the energy sector, particularly the production of biofuels, and combating food contamination in the agricultural industry.
Dr. Gao’s current and future research aspirations in the field of human genetics and the energy sector serve as an inspiration for aspiring female scientists globally. She hopes that young females will follow their passion in STEM, and envisions a future where female scientists are not in the minority. Because of Dr. Gao and her vision, the Gao Laboratory is not only an embodiment of cutting-edge innovation in the future of enzyme-catalyzed reactions, but also the inspirational potential of female scientists.
Works Cited
[1] What are genome editing and CRISPR-Cas9? - Genetics Home Reference - NIH. https://ghr.nlm.nih.gov/primer/genomicresearch/genomeediting (accessed Nov 8, 2019).
[2] Gao, X., Tao, Y., Lamas, V. et al. Nature. [Online] 2018, 553, 217-221. https://www.nature.com/articles/nature25164 (accessed Nov 8, 2019).
[3] Berg, J.; Tymoczco, John; etc. Biochemistry, 5th; W.H Freeman & Co: New York, 2002; 303-341.